红掌钙结合蛋白基因AaCBP1的克隆与低温诱导特性分析 
 作者
作者  通讯作者
通讯作者
《分子植物育种》网络版, 2013 年, 第 11 卷, 第 7 篇 doi: 10.5376/mpb.cn.2013.11.0007
收稿日期: 2013年04月22日 接受日期: 2013年04月24日 发表日期: 2013年05月08日
引用格式(中文):
刘晓静等, 2013, 红掌钙结合蛋白基因AaCBP1的克隆与低温诱导特性分析, 分子植物育种(online), 11(7): 1042-1046 (doi: 10.5376/mpb.cn.2013.11.0007)
引用格式(英文):
Liu et al., 2013, Molecular Cloning of Calcium-binding Protein AaCBP1 Gene and Its Expression Under Cold Stress in Anthurium, Fenzi Zhiwu Yuzhong (online) (Molecular Plant Breeding), 11(7): 1042-1046 (doi: 10.5376/mpb.cn.2013.11.0007)
钙结合蛋白广泛参与植物多种非生物逆境过程。本研究以红掌‘Alabama’为材料,采用PCR法,克隆了1个红掌钙结合蛋白基因AaCBP1(GenBank: KC907407),基因片段长度为663 bp,编码220个氨基酸,具有典型的钙结合保守域。通过氨基酸序列比对结果发现,与红掌AaCBP1同源性较高的是蓖麻的钙结合蛋白,相似度在46%左右。采用RT-PCR方法分析了AaCBP1在低温下的表达模式,结果发现AaCBP1有明显的低温诱导特性,在低温处理3 h表达量就有明显升高。研究表明AaCBP1基因在红掌低温胁迫响应中具有重要作用,可为红掌抗寒分子育种奠定重要基础。
红掌(Anthurium andraeanum)是近年来全球发展较快、需求量较大的高档盆花和切花花卉。红掌原产热带,对温度较为敏感。在生产流通的各个环节中,红掌极易遭受低温冷害,导致植株生长停止,叶片黄化和苞片着色不正常等(田丹青等, 2011),严重影响红掌盆花和切花品质。低温是限制红掌生长发育的重要逆境因子,如何提高红掌的低温耐性已成为红掌生产和流通过程中的迫切需要解决的问题(田丹青等, 2012)。因此,深入研究红掌在低温胁迫下的分子响应机制,对于红掌生产具有十分重要的现实意义和应用价值。
钙结合蛋白是生物体中广泛存在的一类结构保守的调节蛋白。钙结合蛋白通过和Ca2+结合,并与靶蛋白相互作用,调控植物细胞的发育和对外界刺激的反应。研究表明,钙结合蛋白参与多种生理过程,如基因表达,调节细胞分裂和代谢,生殖发育和细胞膜通道调控等。现在已从多种植物,如拟南芥、大麦、水稻、辣椒、甜菜、丹参和芸豆中成功克隆了钙结合蛋白基因。在拟南芥基因组中发现了50多种钙结合蛋白,其序列相似度由 20%~75%不等,这些钙结合蛋白广泛参与了拟南芥的多种生理活动(Tsou et al., 2012; McCormack et al., 2005)。钙离子信号的传导是个复杂的过程,越来越多的钙结合蛋白被发现在植物应答干旱、低温、盐碱等逆境胁迫过程中具有重要作用。在水稻中,CBL基因家族含有10个成员,这10个CBL基因均有6到7个内含子,在水稻逆境防御反应中发挥关键作用(Asano et al., 2002)。在拟南芥中研究表明,CBL1 基因可以响应多种非生物逆境,作为基因网络的上游调控因子,对多种逆境途径的基因转录有调节作用(Albrecht et al., 2003)。
本研究以红掌‘Alabama’为研究材料,克隆了一个红掌钙结合蛋白基因AaCBP1,我们对其核苷酸序列及编码的蛋白特征进行了分析,并对AaCBP1的低温诱导特性进行了研究。本研究初步解析了红掌钙结合蛋白基因的特点和功能,为探索红掌低温胁迫下的分子响应机理打下了良好基础,同时为红掌抗寒分子育种提供了基因资源。
1结果与分析
1.1红掌钙结合蛋白基因片段克隆
经过红掌EST序列的大规模测序,发现一个受冷胁迫诱导的钙结合蛋白,设计特异引物,多次测序确定了该基因序列。获得的基因片段长度为663 bp,编码220个氨基酸(图1) (GenBank登录号:KC907407)。
 图1红掌钙结合蛋白基因核苷酸序列及编码的氨基酸序列 Figure 1 Anthurium calcium-binding protein gene nucleotide sequence and amino acid sequence |
1.2红掌AaCBP1基因片段序列分析
将获得的基因片段在Genebank数据库中进行BlastX比对,发现该基因与拟南芥、蓖麻、玉米、水稻、芸豆、胡杨等的钙结合蛋白具有较高的同源性。氨基酸序列比对结果发现,同源性较高的是蓖麻(Ricinuscommunis)的钙结合蛋白,相似度在46%左右(图2)。在NCBI进行保守域结构预测发现,结果显示AaCBP1编码的蛋白有一个植物保守的钙结合结构域,具有该结构域的蛋白可能参与植物多种防御响应(图3)。我们把该基因命名为AaCBP1。采用DNAMAN对已知物种的钙结合蛋白序列进行比对和系统发育树分析,结果显示红掌AaCBP1蛋白与芸豆的钙结合蛋白亲缘分支关系较近(图4)。
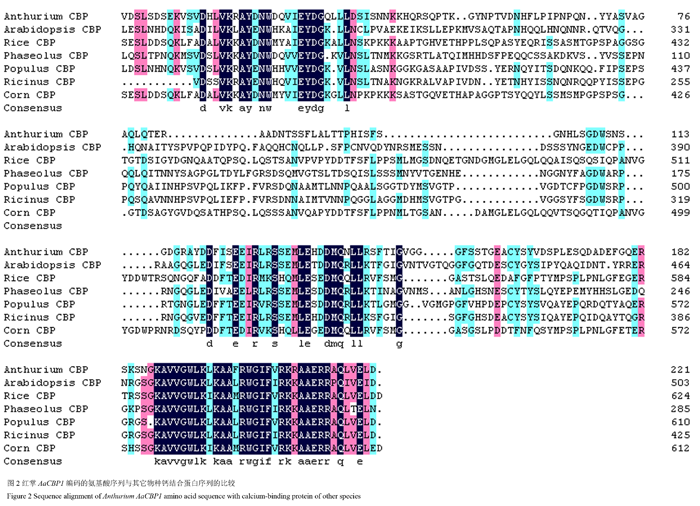 图2 红掌AaCBP1编码的氨基酸序列与其它物种钙结合蛋白序列的比较 Figure 2 Sequence alignment of Anthurium AaCBP1 amino acid sequence with calcium-binding protein of other species |
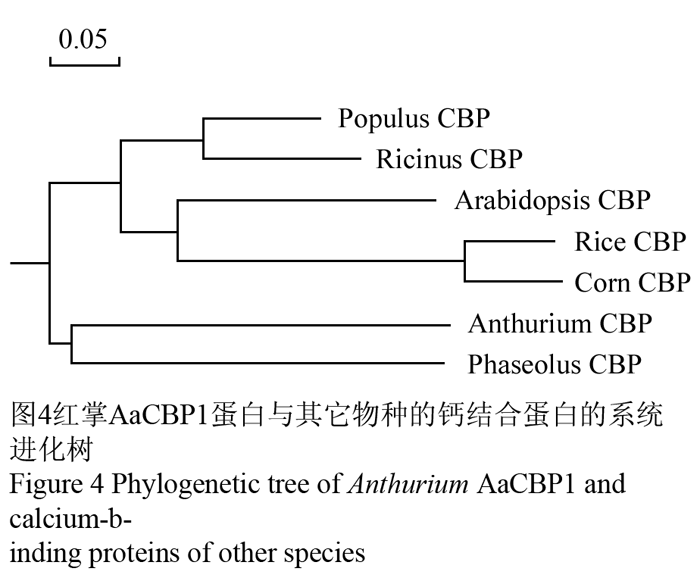 图4红掌AaCBP1蛋白与其它物种的钙结合蛋白的系统进化树 Figure 4 Phylogenetic tree of Anthurium AaCBP1 and calcium-binding proteins of other species |
1.3红掌AaCBP1基因低温诱导特性分析
从红掌冷处理和对照样本中分别提取叶片总RNA,反转录合成cDNA。以获得的cDNA为模板,RT-PCR 检测AaCBP1在低温下的表达模式。从图5可以看出,AaCBP1基因表达受低温诱导,在低温处理3 h表达量就有明显升高。而在对照植株中该基因表达稳定,不存在诱导表达。结果表明AaCBP1基因可能在红掌的低温响应过程中起重要作用。
|
图5 RT-PCR分析红掌AaCBP1基因的低温诱导特性 Figure 5 RT-PCR analysis of induction of Anthurium AaCBP1 gene expression under low temperature |
2讨论
低温等逆境是植物生长、发育的主要环境限制因素,植物经过长期生存演化形成一套感知低温信号的精细调控机制。钙结合蛋白在植物的信号转导过程中起关键作用,其相关机理的研究一直是植物生理学和细胞生物学研究的热点(毛国红等, 2004)。在拟南芥中,钙结合蛋白CBL1在干早胁迫诱导下,基因的表达量上升,但在冷刺激下,基因的表达量却受到抑制,CBL1基因在低温胁迫下具有负调控作用(Albrecht et al., 2003)。在玉米中,10个CBL基因的低温响应特点不同,有的为诱导表达,有的则为抑制表达(Yang and Poovaiah, 2003)。在红掌中,我们克隆得到的AaCBP1基因为低温诱导表达,其对红掌的低温胁迫响应可能具有正调控作用。由于钙结合蛋白在植物中为多基因家族,在红掌中还可能存在多个钙结合蛋白,共同调控红掌的低温响应。
钙结合蛋白不仅参与植物低温胁迫响应,还在植物生长发育和多种防御反应中发挥重要作用。在拟南芥中发现,CMLs与植株的病原菌应激反应,盐胁迫等有关(Chiasson et al., 2005; Yamaguchi et al., 2005)。钙结合蛋白PCaP2参与了拟南芥根毛的发育过程(Kato et al., 2013)。在水稻和萝卜中研究发现,钙结合蛋白在植物胚轴生长和和细胞分裂等生长发育过程中有重要作用(Cocucci and Negrini, 1988; Asano et al., 2002)。红掌钙结合蛋白AaCBP1参与了低温胁迫响应,其在红掌生长发育和其它逆境胁迫中的作用尚不明确,因此,深入研究红掌钙结合蛋白功能具有深远意义和较高应用价值。
3材料与方法
3.1材料
红掌(Anthurium andraeanum) ‘Alabama’幼苗生长于浙江省农科院花卉研究开发中心温室中。大肠杆菌DH5α为浙江省农科院花卉研究开发中心实验室保存。
3.2 RNA提取、基因克隆
取红掌叶片6℃低温胁迫下0, 3 h,6 h,12 h,24 h 的样本,利用Trizol (Invitrogen, USA)法提取叶片的Total RNA。根据该试剂的说明书进行操作。紫外分光光度计测定总RNA 230 nm、260 nm、280 nm和300 nm吸收值,计算总RNA含量,估测其纯度。然后将RNA样本保存于-80℃超低温冰箱中备用。
利用已知的基因序列,设计特异引物,克隆钙结合蛋白基因片段,把克隆得到的片段连接到T-easy载体,转入大肠杆菌DH5α,经多次测序确定基因序列。利用NCBI、DNAMAN等软件对核苷酸序列和编码的氨基酸序列进行分析。
3.3 RT-PCR分析基因表达模式
采用RT-PCR分析基因的表达模式,所用引物序列见表1。从红掌冷处理和对照样本中分别提取总RNA,取1 μg总RNA用来做反转录(Powerscript reverse transcriptase, Clontech, CA, USA)。PCR扩增条件为:94℃预变性4 min,随后为特定循环数的94℃变性30 s,57℃退火30 s,72℃延伸45 s。其中,AaCBP1设27个循环,Actin设25个循环。最后一个循环结束后,72℃再延伸5 min。PCR产物用1.2 % (w/v)的琼脂糖凝胶进行分离,EtBr染色分析。
作者贡献
刘晓静是本研究的实验设计和实验研究的执行人;刘晓静、田丹青和及潘晓韵完成数据分析,论文初稿的写作;葛亚英,王炜勇和潘刚敏参与实验设计,试验结果分析;郁永明是项目的构思者及负责人,指导实验设计,数据分析,论文写作与修改。全体作者都阅读并同意最终的文本。
致谢
本研究由浙江省自然科学基金(LQ12C15003)资助。
参考文献
Albrecht V., Weinl S., Blazevic D., D'Angelo C., Batistic O., Kolukisaoglu U., Bock R., Schulz B., Harter K., and Kudla J., 2003, The calcium sensor CBL1 integrates plant responses to abiotic stresses, Plant J., 36(4): 457-470
http://dx.doi.org/10.1046/j.1365-313X.2003.01892.x
PMid:14617077
Asano T., Kunieda N., Omura Y., Ibe H., Kawasaki T., Takano M., Sato M., Furuhashi H., Mujin T., Takaiwa F., Wu C.Y., Tada Y.C., Satozawa T., Sakamoto M., and Shimada H., 2002, Rice SPK, a calmodulin-like domain protein kinase, is required for storage product accumulation during seed development: phosphorylation of sucrose synthase is a possible factor, Plant Cell, 14(3): 619-628
http://dx.doi.org/10.1105/tpc.010454
PMid:11910009 PMCid:150584
Chiasson D., Ekengren S.K., Martin G.B., Dobney S.L., and Snedden W.A., 2005, Calmodulin-like proteins from Arabidopsis and tomato are involved in host defense against Pseudomonas syringaepv.tomato, Plant Mol. Biol., 58(6): 887-897
http://dx.doi.org/10.1007/s11103-005-8395-x
PMid:16240180
Cocucci M., and Negrini N., 1988, Changes in the levels of calmodulin and of a calmodulin inhibitor in the early phases of radish (Raphanussativus L.) seed germination -effects of ABA and fusicoccin, Plant Physiol., 88(3): 910-914
http://dx.doi.org/10.1104/pp.88.3.910
PMid:16666403 PMCid:1055681
Kato M., Aoyama T., and Maeshima M., 2013, The Ca2+-binding protein PCaP2 located on the plasma membrane is involved in root hair development as a possible signal transducer, Plant J., doi: 10.1111/tpj.12155
http://dx.doi.org/10.1111/tpj.12155
Mao G.H., Song L.X., and Sun D.Y., Progress of study on calmodulin-binding proteins in plants, Zhiwu Shengli Yu Fenzi Shengwu xue Xuebao (Journal of Plant Physiology and Molecular Biology), 30(5): 481-488 (毛国红, 宋林霞, 孙大业, 2004, 植物钙调素结合蛋白研究进展, 植物生理与分子生物学学报, 30(5): 481-488)
McCormack E., Tsai Y.C., and Braam J., 2005, Handling calcium signaling: Arabidopsis CaMs and CMLs, Trends Plant Sci., 10(8): 383-389
http://dx.doi.org/10.1016/j.tplants.2005.07.001
PMid:16023399
Tian D.Q., Ge Y.Y., Pan G.M., Liu X.J., Shen X.L., and Pan X.Y., 2012, Effect of different exogenous substances on cold resistance of Anthurium, Zhejiang Nongye Kexue (Journal of Zhejiang Agricultural Sciences), (8): 1142-1144 (田丹青, 葛亚英, 潘刚敏, 刘晓静, 沈晓岚, 潘晓韵, 2012, 不同外源物质处理对红掌抗寒性的影响, 浙江农业科学, (8): 1142-1144)
Tian D.Q., Ge Y.Y., Pan G.M., Shen X.L., He Y., Zhang Z., Liu X.J., Zhu Z.J., 2011, Morphological and physiological characteristics of different cultivars of Anthurium andraenum under chilling stress, Yuanyi Xuebao (Acta Horticulturae Sinica), 38(6): 1173-1179 (田丹青, 葛亚英, 潘刚敏, 沈晓岚, 何勇, 张智, 刘晓静, 朱祝军, 2011, 低温胁迫对3个红掌品种叶片形态和生理特性的影响, 园艺学报, 38(6): 1173-1179)
Tsou P.L., Lee S.Y., Allen N.S., Winter-Sederoff H., and Robertson D., 2012, An ER-targeted calcium-binding peptide confers salt and drought tolerance mediated by CIPK6 in Arabidopsis, Planta, 235(3): 539-552
http://dx.doi.org/10.1007/s00425-011-1522-9
PMid:21971994
Yamaguchi T., Aharon G.S., Sottosanto J.B., and Blumwald E., 2005, Vacuolar Na+/H+ antiportercation selectivity is regulated by calmodulin from within the vacuole in a Ca2+- and pH-dependent manner, Proc. Natl. Acad. Sci., USA, 102(44): 16107-16112
http://dx.doi.org/10.1073/pnas.0504437102
PMid:16249341 PMCid:1276053
Yang T.B., and Poovaiah B.W., 2003, Calcium/calmodulin-me-
diated signal network in plants, Trends Plant Sci., 8(10): 505-512
http://dx.doi.org/10.1016/j.tplants.2003.09.004
PMid:14557048